CAPITULO 1
APOPROTEINAS
Apoproteinas
Lipoproteinas
Metabolismo das lipoproteinas
Metabolismo das apo B-100
Receptores
Dislipemias
Hipolipemias
Bibliografia
Lipoproteinas
Metabolismo das lipoproteinas
Metabolismo das apo B-100
Receptores
Dislipemias
Hipolipemias
Bibliografia
Conceito
Os lipidos, excepto os fosfolipidos são insolúveis na água. Para circularem num meio aquoso como o sangue têm que ser solubilizados. Tal objectivo é conseguido envolvendo-os com moléculas tendo um polo solúvel nos lípidos e outro na água, orientando-se o polo liposolúvel para o interior e o hidrosolúvel para o exterior. As únicas moléculas que têm capacidade para exercer esta função são as proteinas . Os lípidos combinados com proteinas denominam-se lipoproteínas e as proteínas combinadas com os lipidos apolipoproteínas ou simplesmente apoproteínas.

cortesia do prof. William Christie
Segundo Alaupovic uma proteína só poderia ser considerada apoproteina desde que preenchesse os seguintes critérios:
· Ter propriedades únicas e distintas sob os pontos de vista químico, físico e imunológico.
· Ter propriedades distintas nos domínios estrutural e/ou funcional.
· Ser um componente integral do sistema de transporte de lípidos.
· Terem capacidade para formar lipoproteinas.
Para uniformizar a designação das apoproteinas, Alaupovic propoz o sistema ABC, em que as apoproteionas se designam por letras maiúsculas: A,B,C…
Polipeptidos constituintes das proteinas
Algumas proteinas não têm uma cadeia única mas são constituídas por vários polipéptidos. Para Alaupovic estes polipeptidos devem preencher os seguintes critérios:
· Terem propriedades únicas e distintas
· Serem componente integral do sistema de transporte de lípidos
· Participar com outros polipeptidos na composição de uma apoproteina
Os polipeptidos designam-se acrescentando números romanos à apoproteína: A-I, C-III….
Variantes dos polipeptidos
Alguns polipeptidos podem ter variantes. Estas designam-se pelo acréscimo de um número arabe: AI.1, CII.2…
Famílias lipoproteicas
Alaupovic postulou que as lipoproteinas do plasma são constituídas por famílias lipoproteicas.
Uma família lipoproteica é um sistema polidisperso de partículas contendo apenas uma apoproteina.Quando a lipoproteina tem apenas uma familia, ou seja, uma única apoproteina esta diz-se primária, simples ou discreta. Quando a lipoproteina é constituida por mais de uma familia, ou seja, por mais de uma apoproteina esta diz-se secundária ou complexa.
Segundo este conceito as lipoproteinas designar-se-iam pelas apoproteinas constituintes

Tipos de apoproteinas
Descreveram-se várias apoproteinas.
Apoprotein
|
Molecular weight
|
Lipoprotein
|
Function
|
Apo A1
|
28,100
|
HDL
|
Lecithin:cholesterol acyltransferase (LCAT) activation. Main structural protein.
|
Apo A2
|
17,400
|
HDL
|
Enhances hepatic lipase activity
|
Apo A4
|
46,000
|
CM
|
Apoproteins.*
|
Apo B48
|
241,000
|
CM
| |
Apo B100
|
512,000
|
LDL, VLDL
|
Binds to LDL receptor
|
Apo C1
|
7,600
|
VLDL, CM
|
Activates LCAT
|
Apo C2
|
8,900
|
VLDL, CM
|
Activates lipoprotein lipase
|
Apo C3
|
8,700
|
VLDL, CM
|
Inhibits lipoprotein lipase
|
Apo D
|
33,000
|
HDL
|
Associated with LCAT, progesterone binding
|
Apo E
|
34,000
|
HDL
|
At least 3 forms. Binds to LDL receptor
|
Apo(a)
|
300,000-800,000
|
LDL, Lp(a)
|
Linked by disulfide bond to apo B100 and similar to plasminogen
|
Apo H, J, L, M
|
Poorly defined functions
| ||
* Roman numerals are sometimes used to designate apoproteins (e.g. apo AI, AII, AIII, etc)
| |||
Cortesia do prof. William Christie
Estrutura e funções das diferentes apoproteinas
Apoproteina A
Descreveram-se quatro polipeptidos:
Apo A-I
Encontra-se especialmente nas HDL quer isolada (forma primária) quer associada à A-II (forma secundária).
É constituída por 243 aminoácidos com seis segmentos muito homologos. Há varias variantes designadas pelo nome das cidades onde foram descobertas: AI-Milão, AI-Marburgo…
É sintetizada no fígado e no intestino como pro-apo A-I que na circulação se transforma em apo A-I.
A sua principal função consiste em combinar-se com receptores celulares específicos para captar colesterol das membranas celulares e efectuar o efluxo do colesterol. Esta acção é especifica das A-I pois a lipoproteina AII é muito fracamente captada pelos receptores ao contrário da AI:AII. Além disso a AI é um activador da LCAT (Lecitina Colesterol Acil Transferase) enzima responsável pela esterificação do colesterol.
Apo A-II
Contém dois polipeptidos iguais de 77 aminoacidos ligados por uma ponte S-S na cisteina 6.
É sintetizada no figado como uma preapoproteina de 100 aminoacidos
O seu significado fisiológico ainda não está compreendido
Apo A-Encontra-se principalmente nas HDL. É constituída por dois polipetidos iguais, cada um com 73 aminoácidos, ligados entre si por uma ponte S-S.
A sua função não é bem conhecida. Pode ligar-se a fosfolípidos para formar partículas solúveis. In vitro inibe a LCAT mas parece que in vivo esta acção depende da razão A-I/A-II.
Apo A-IV
É o maior componente das HDL nos ratos mas no homem é um componente minor.
A sua função é desconhecida
Apoproteina B
Encontra-se nas LDL, VLDL, IDL e quilomicra. Enquanto que nas HDL é a única apoproteina, nas outras lipoproteinas encontra-se associada.
Kane caracterizou várias formas de apo B com pesos moleculares diferentes. À proteina com maior peso molecular deu o nome de apo B-100 (100=100%). As outras têm um número corresponde à fracção correspondente da 100 (B-74, B-48, B-37, B-26): As únicas que têm interesse biológico são a B-100 e a B-48.
A apo B-100 é um polipeptido de 4500 aminoacidos com varias unidades de repetição unidas por ligações covalentes. É sintetisada no fígado. Encontra-se como apoproteina única das LDL, encontrando-se também das VLDL, LDL, IDL e Lp(a).
É a proteina determinante para o reconhecimento das LDL pelos receptores celulares.
A apo B-48 é sintetisada no intestino. É a proteina estrutural dos quilomicra, não se encontrando nas LDL.
As apoB-100 e B-48 provêm do mesmo gene mas o m-RNA intestinal é modificado pela formação de um codão stop Não se conhece o enzima que opera esta transformação, mas tem uma especitidade de orgão muito elevado
Apoproteina C
Descreveram-se três polipeptidos: C-I, C-II e C-III
Apo C-I
60% encontram-se nas HDL e 40% nas VLDL, associada a outras proteínas.
É uma cadeia simples de 57 aminoácidos sendo rica em aminoácidos básicos ( 9 moléculas de lisina e 3 de arginina). Desconhece-se o seu local de síntese e há poucos dados sobre o seu papel fisiológico embora in vitro iniba a lipoproteina lipase e active a LCAT.
Apo C-II
É uma cadeia simples de 79 aminoácidos, estando o seu gene localizado no cromosoma 19, sendo sintetizada no figado.
É o cofactor da lipoproteina lipase sendo portanto um activador potente deste enzima.
Apo C-III
É a mais abundante das apoproteinas C. Tem 79 aminoácidos contendo uma molécula de galactose e outra de galactosamina. O seu conteúdo em ácido siálico define a existência de três variantes
.Variantes da apo CIII
Variante Moléculas de ac. siálico
.
CIII:0 0
CIII:1 1
CIII:2 2
O seu gene está colocado no cromosoma 11 na extremidade 3 do gene da A-I.
Inibe a interacção proteina-receptor impedindo por consequência o catabolismo das LDL mediado por receptores .Por outro lado inibe in vitro a lipoproteina lipase com a consequente diminuição da depuração dos quilomicra.
Apoproteina D
Encontra-se isolada ou associada com outras apoproteinas nas HDL. A sua estrutura não é conhecida. Contem l8% de glúcidos sendo a heterogeneidade desta acção que explica a existência de seis isoformas.As três isoformas mais frequentes são determinados por locus separados - e4,e 3 e e2) originando seis fenotipos.
Polimorfismo das apo E

Há ainda variantes raras . A maior parte destas variantes está associada com a dislipémia familiar.
Uma função totalmente diferente das apo e é a sua presença no sistema nervoso central. O cérebro é a seguir ao figado o orgão que contém mais mRNA da apo E.
A apo E é necessaria para o crescimento e regeneração do sistema nervoso central e a sua síntese aumenta após uma lesão. A apo E também se encontra aumentada em algumas doenças crónico-degenerativas. Na doença de Alzheimer parece estar associada com o alelo e4.
Apoproteina F
É a apoproteína de natureza mais ácida. É uma proteina minor das HDL.
Apoproteina G
É uma proteína minor das HDL
Apoproteina H
É uma apoproteína minor sem função conhecida. Descreveu-se a sua ausência em cinco individuos sem existir qualquer alteração do metabolismo lipidico.
Apoproteina I
Malmendier e col isolaram das HDL uma outra apoproteína a que denominaram inicialmente de apoproteina S (S=sugar induced) por ser induzida pela ingestão de açucar. Posteriormente, para seguir a nomenclatura de Alaupovic estes autores propuseram a sua designação como apoproteina I.
Apoproteina J ou cluterina
É uma proteina de 4277 aminoácidos cindida após a tradução em duas subunidades ligadas por uma ponte S-S.
Existe nas HDL e pode associar-se ao colesterol e fosfolípidos para formar complexos imunocirculantes.
Funções das apoproteinas
Do que dissemos neste capitulo poderemos constituir que se pode atribuir às apoprotrinas as seguintes funções
· Componentes das lipoproteínas
· Transporte transcelular de lípidos
1. Transporte do colesterol (apo B)
2. Catabolismo mediado por receptores (apos B e E)
3. Efluxo do colesterol (Apo A-I)
· Cofactores de enzimas
· Inibidores de enzimas
Funções das apoproteinas

Das apoproteinas às lipoproteinas
Para uma proteína originar uma lipoproteina esta terá que ser sintetizada, sofrer modificações intra e extracelulares, incorporar os lípidos correspondentes e sofrer remodelação e transferência no plasma.
Síntese
Nos mamíferos as proteinas são sintetizadas quasi exclusivamente no fígado e intestino, exceptuando a apoproteina E que também é sintetizada no cérebro. Alguns orgãos têm uma produção minor de apoproteinas .
Locais major de síntese das apoproteinas0
Apoproteinas Locais de síntese
AI fígado, intestino
AII fígado
B100 figado, intestino
B48 intestino
CI fígado, suprarenais
CII e CIII fígado
E fígado, cérebro,rins, suprarenais
Modificações intracelulares
Propeptidos
A maior parte das apoproteinas são sintetizadas com um peptido terminal de 16 a 25 aminoacidos que será cindido por uma peptidase da membrana (peptidase sinal)ou do retículo.
Fosforilação
A apoproteina B é segregada fosforilada. Em culturas a insulina diminui a fosforilação facilitando a sua degradação intracelular.
Glicosilação
No aparelho de Golgi algumas apoproteinas (B, CIII, D, E) podem ser glicosiladas, tendo as cadeias glucídicas ácido siálico terminal.
Sialilação
As apo CIII têm variantes não sialiladas (CIII:0) e sialiladas (CIII:1 e CIII:2).
Modificações extracelulares
Preproapoproteinas
A apo apo A-I é sintetizada como uma preapoproteina. A peptidase sinal cinde-a em prepeptido e proapoproteina que após ser segregada é cindida por uma protease em apoproteina e propeptido .

Dessialação
Todas proteínas sialiladas são desialiladas após a sua secreção

Outras modificações
Na diabetes pode ocorrer uma glicosilação não enzimática
Também foram descritas desamidação e transglutaminação de apoproteinas
Incorporação de lipidos
Apo B
A apo B associa-se à membrana do retículo, transferindo-se esta apo B ligada à membrana para o retículo onde se combina com os triglicéridos. Os fosfolípidos e colesterol são incorporados no aparelho de Golgi.
Outras apoproteinas
As apoproteinas A-I.E e CIII são segregadas numa forma pobre em lípidos. Para a incorporação intracelular todas as apoproteinas necessitam de apo B.
A incorporação pode ser reformulada na circulação sistémica. A apo E pode ser incorporada em lipoproteinas após atingir a circulação sistémica.
Remodelagem e transferência
Após a sua secreção sofrem alterações do plasma. Esta remodelagem baseia-se nos princípios seguintes:
· As apoproteinas segregadas originam lipoproteinas nativas que no plasma se transformarão em lipoproteinas maduras.
· A maturação das lipoproteinas faz-se no compartimento intravascular por um processo de remodelagem afectando essencialmente a redistribuição de apoproteinas
· Na sua remodelagem ou na sua sintese de novo as lipoproteinas adquirem as apoproteinas necessárias para a sua função
Capitulo 2
LIPOPROTEINAS
Classificação
Há várias proteínas que diferem entre si pela natureza dos lipidos transportados e pelas proteinas que contêm. A ignorância sobre a sua estrutura exacta levou a classificá-las conforme os seus métodos de separação mais usuais.
Ultracentrifugação
Gofman constatou que ultracentrifugando o soro a uma densidade de l063 havia uma partícula mais pesada que sedimentava e outra mais leve que flutuava. Recentrifugando o sobrenadante a uma densidade de1006 observou uma que sedimentava e outra que flutuava .
Verificou assim existência de três classes de lipoproteinas conforme a sua densidade:

A estas classes deve-se ainda acrescentas os quilomicras mais leves, flutuando no soro sem ultracentrifugação.
A densidade das lipoproteinas é inversamente proporcional ao seu diâmetro.
Electroforese
A utilização da electroforese de zona para a separação das lipoproteinas foi efectuada em l941 mas só a introdução da electroforese em papel em 1955 perm.itiu a sua vulgarização. As lipoproteinas foram classificadas pela sua ordem de migração:
- · Lipoproteinas sem mobilidade electroforetica - quilomicra
- · Lipoproteinas com mobilidade semelhante às a-globulinas - a - lipoproteinas
- · Lipoproteinas com mobilidade b-globulina (b-lipoproteinas).
- · Lipoproteinas com motilidade a2 - pre-b-liproteinas
Equivalencia entre os dois sistemas

Estudo individual das lipoproteinas
Quilomicra
Este nome foi utilizado pela primeira vez em 1920 para descrever proteínas visíiveis ao microscópio óptico que aparecem no plasma após uma refeição gorda. Esta visibilidade deve-se às suas grandes dimensões (75 a 600 nm)
São particularmente ricos em triglicéridos (80-90%) o que explica a sua baixa densidade. Os outros componentes são colesterol livre (1-3%), colesterol esterificado (3-6), fosfolípidos (7-9) e apoproteinas (1-2) .
Os quilomicra nativos têm apo A-I (apoproteina major), apo B48 (sintetizada pelas proteínas intestinais), apo A-IV e apo E. Ao chegarem à circulação sistémica enriquecem-se com apo C-II
{kind=link}

{kind=link}
VLDL.
São moléculas grandes embora de dimensões inferiores às dos quilomicra. Não interceptam os feixes luminosos e por isso não são visíveis ao microscópio óptico mas quando em quantidade dão um aspecto turvo ao soro.
São a seguir aos quilomicra as lipoproteinas mais ricas em triglicéridos . Os outros constituintes são colesterol livre (6-8), colesterol esterificado (10-15), fosfolípidos (17-18) e apoproteinas (9-10).
As apoproteinas major são apo B -100 e Apo C, encontrando-se a Apo E em pequenas quantidades.
LDL
Em contraste com a riqueza dos quilomicra e VLDL em triglicéridos, as LDL são particularmente ricas em colesterol (-46-5O%), estando a maior parte esterificada (80-87%). Os outros constituintes são trigliceridos (5-8), fosfolipidos (20-25) e apoproteinas (20-24) . 95% das apoproteinas são representadas pela apo B-1000) havendo pequenas quantidades de apo C e apo E e quantidades vestigiais de A-I e A-II

cortesia do prof. Foster
HDL
São as lipoproteinas mais pequenas e mais densas devido ao seu alto conteúdo em apoproteinas (cerca de 50%), sendo a predominante a Apo A-I (70-80% das proteínas totais) encontrando-se ainda a apo A-II (20-23) e C (5-13) encontrando-se em quantidades menores as D, E e A-IV.
Separaram-se duas subclasses, as HDL2 e HDL3. As HDL2 são maiores e mais densas tendo mais lípidos e menos apoproteinas que as HDL2.
Lipoproteinas
Density (g/mL)
|
Class
|
Diameter (nm)
|
% protein
|
% cholesterol
|
% phospholipid
|
% triacylglycerol
|
>1.063
|
5-15
|
33
|
30
|
29
|
8
| |
1.019-1.063
|
18-28
|
25
|
50
|
21
|
4
| |
1.006-1.019
|
IDL
|
25-50
|
18
|
29
|
22
|
31
|
0.95-1.006
|
30-80
|
10
|
22
|
18
|
50
| |
<0.95
|
100-1000
|
<2
|
8
|
7
|
84
|

Capitulo 3
METABOLISMO DAS LIPOPROTEINAS
O metabolismo das lipoproteinas é o reflexo das apoproteinas que as constituem. É portanto lógico estudar o seu metabolismo integrado no das apoproteinas.
Digestão dos lipidos
Esteres do colesterol
Os esteres do colesterol são hidrolisados pela colesterol esterase em colesterol e ácidos gordos . O colesterol é absorvido pelas células intestinas na presença de ácidos gordos

Fosfolipidos
As fosfolipases hidrolisam os fosfolipidos originando uma molécula com menos um ácido gordo (lisolecitina no caso da lecitina) e ácidos gordos

Triglicéridos
{kind=link}
No intestino a lipase pancreática hidrolisa progressivamente os triglicéridos com formação progressiva de ácidos gordos, di e monogliceridos

cortesia de Dick Bowen
Destino dos ácidos gordos
Os ácidos gordos libertados terão destinos diferentes conforme o comprimento da sua cadeia
Ácidos gordos de curta e média cadeia
Estes ácidos têm um numero de atomos de carbono inferior a 12 e representam uma percentagem reduzida da totalidade dos ácidos gordos. Não são absorvidos pela célula intestinal, entrando directamente para a veia porta
Ácidos de longa cadeia
São absorvidos pela célula intestinal, necessitando estar sob a forma de micelas, para o que são necessarios ácidos biliares

Destino dos acidos gordos

Micelas
Formação de quilomicra
Os acidos gordos são transformados no retículo liso da célula intestinal em triglicéridos, combinando-se com apoproteinas para formar quilomicra

Embora a Apo A seja a apoproteina major dos quilomicra apo B-48 é necessária para a formação dos quilomicra. Alguns minutos após a ingestão de gorduras há aumento da síntese de apo B-48. Por outro lado na a-b-lipoproteinemia (doença em que falta apo B) não se formam quilomicra.

Maturação dos quilomicra
Os quilomicra nativos são ricos em triglicéridos, tendo pouco colesterol. A maturação começa na linfa onde recebem colesterol livre e algumas apoproteinas, mas as modificações mais importantes fazem-se na circulação sistémica:
Aquisição de apo C proveniente de outras lipoproteínas
As apoproteinas provêm em especial das HDL . Esta aquisição modula a lipólise pois que a apo C-II é o cofactor da lipoproteina lipase
Hidrólise dos trigliceridos pela lipoproteina lipase
As partículas são sequestradas no endotélio por fusão com os fosfolípidos da membrana externa da membrana, dando assim a possibilidade de o enzima actuar sobre os triglicéridos no interior da molécula. Como consequência da hidrólise liberta-se apoproteina A-I, utilizada na formação de HDL2 a partir das HDL3 e alguma apo C.
Devido a esta intervenção formam-se os remanescentes dos quilomicra, partículas pobres em trigliceridos e ricas em colesterol

Formação dos quilomicra

Aquisição de apo E pelos remanescentes
O enriquecimento em apo E permite o reconhecimento destas particulas pelos receptores pois estes não reconhecem a apo B-48
Captação dos remanescentes
Cerca de 25 a 30% dos remanescentes são captados pelos receptores BE. Admite-se a existência de dois outros receptores para a apo E - os receptores E e a LRP (LDL receptor related protein).
Catabolismo dos quilomicra na medula óssea
Embora a maior parte do catabolismo se faça no figado, estes tambem são metabolizados na medula óssea, tendo-se-lhe atribuído os significados seguintes:
· Fornecimento de lipidos como fonte de energia para a síntese da membrana durante a hematopoiese
· Manutenção de um nível adequado de lipidos - O turnover de ácidos gordos nos adipocitos da medula é muito maior que nos outros adipocitos. Os triglicéridos são utilizados como fonte de energia durante a proliferação celular e há uma relação inversa entre o conteudo nos adipocitos e a hematopoiese
· Libertação de vitaminas liposoluveis como a vitamina A
Metabolismo da apo B-100
Formação das VLDL
A sua formação é muito semelhante à dos quilomicra. Os ácidos gordos libertados por hidrolise dos trigliceridos ou sofrem b-oxidação ou são armazenados como triglicéridos. Os ácidos gordos seriam captados pelos tecidos extra-hepáticos (músculo, tecido adiposo, coração) e numa menor extensão pelo fígado e resintetizados em triglicéridos no figado. Os triglicéridos dirigem-se para o retículo liso e depois para o aparelho de Golgii onde se combinam com a apoproteina B-100 sintetizada no reticulo rugoso para formar vesiculas de VLDL nativas que migram para a membrana basal.
As VLDL deixam as vesículas por exocitose indo para a circulação sistémica através dos espaços de Disse .
Ácidos gordos
Triglicéridos
Retículo liso
GOLGI
__________________________________
Apo B100
Retículo rugoso
Vesículas de VLDL nativas
Exocitose
Espaços de Disse
Circulação
Maturação das VLDL
As VLDL nativas tornam-se maduras pelo enriquecimento em colesterol devido à acção da LCAT e por transferência das HDL


Formação dos remanescentes das VLDL
A lipoproteina lipase hidrolisa os triglicéridos ao mesmo tempo que toda a apo A e grande parte da apo C é transferida para as HDL, recebendo em troca apo E
No decorrer destas trocas a apo B-100 não é trocada pelo que os remanescentes das VLDL assim formados têm pouca apo A e apo C mas são ricos em apo B-100 e apo E ao mesmo tempo que são pobres em trigliceridos e ricas em colesterol esterificado.

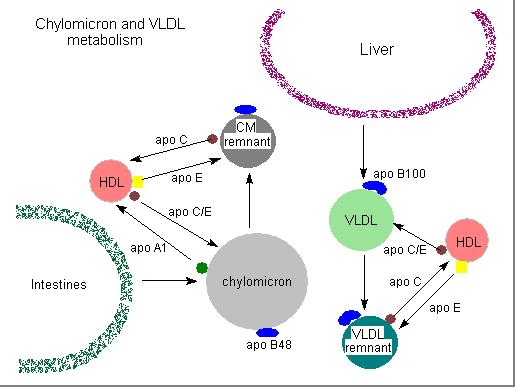
cortesia do prof. William Christie
Estes remanescentes são geralmente descritos como IDL e b-VLDL. Para a maior parte dos autores os remanescentes habitualmente formados são as IDL, formando-se as b-VLDL apenas após uma alimentação rica em colesterol.
Destino dos remanescentes das VLDL
Na maior parte dos animais os remanescentes são reconhecidos pelos receptores BE do figado . No homem só 50% têm este destino. Os remannescentes não reconhecidos são captados pelos receptores mas não internalisados , sendo os trigliceridos hidrolisados pela triglicerido lipase hepática formando-se as LDL nativas
Maturação das LDL
As LDL nativas perdem apo E e recebem colesterol para se transformarem na sua forma madura, muito rica em colesterol e contendo apenas apo B.
Catabolismo da apo B-100
Será estudado com os receptores:
Semelhança entre o metabolismo dos quilomicra e das VLDL
Há alguns pontos comuns entre o metabolismo destas duas lipoproteinas
· Transporte de trigliceridos. Os quilomicra transporam trigliceridos alimentares (trigliceridos exogenos) e as VLDL trigliceridos de origem metabolica (Trigliceridos endogenos)
· Necessidade da apo B para a sua formação, B48 no intestino e B-100 no figado
· Analogia na degradação . Tanto os quilomicra como as VLDL transformama-se em remanescentes - remanescentes dos quilomicra no primeiro caso, IDL e b-VLDL no segundo
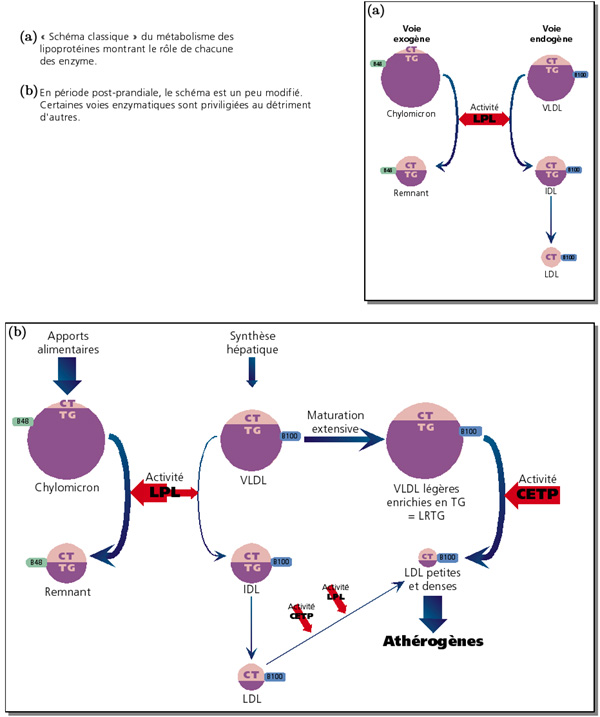
cortesia de J.M.Botto
Metabolismo da apoproteina A
Transporte reverso do colesterol
As lipoproteinas contendo apo B têm a função de transportar lipidos para as células.
As contendo apo A transportam colesterol das celulas para o fígado para aí ser catabolisado - é o transporte reverso do colesterol

Este transporte reverso faz-se em três fases
· Efluxo do colesterol das celulas exta-hepaticas para o plasma.
· Modificações intravasculares do colesterol por um processo envolvendo a LCAT e a transferência dos esteres do colesterol para a apoproteina B.
· Transferência do colesterol para as células hepáticas.

Efluxo do colesterol
O efluxo do colesterol é determinado pela composição da membrana celular e das lipoproteinas aceitadoras
Composição da membrana
A natureza dos microdominios membranários envolvidos no efluxo do colesterol não é bem conhecida. O local mais provável parece ser representado por invaginações da membrana sem clatrina (caleolae) caracterisadas pela presença de caveolina. Nas caveolae encontra-se mais colesterol livre e fosfolípidos que nas outras partes da membrana. As caveolae tambem contêm proteina G provavelmente envolvidas na transdução do sinal induzido pelas HDL
Lipoproteinas aceitadoras
A eficiência do efluxo do colesterol depende também de diferenças no tamanho, fluidez e composição proteica das lipoproteinas aceitadoras
· Tamanho. As particulas pequenas são mais efectivas que as grandes. Descreveram-se conforme as dimensões três subclasses de LpA-I, sendo as de dimensão menor muito mais efectivas.
· Fluidez. O efluxo de colesterol aumenta com a fluidez das partíiculas. A insaturação e a menor dimensão das cadeias dos ácidos gordos aumentam a fluidez
· Composição. A apoproteina A-I é muito mais efectiva que a A-II e facilita o efluxo.
Modificações intravasculares
A transição estrutural das HDL nativas, discoides, em esteres do colesterol e trigliceridos, para HDL maduras, esféricas, com um núcleo central de esteres do colesterol é catalisada pela LCAT. No decurso desta conversão a apo E é substituída por apo A-II até a relação E/AI passar para 1/7.
O aumento de hidrofobia provocado pela esterificação do colesterol leva os esteres a moverem-se para o interior do disco originando a forma esferica e deixando na periferia espaço necessário para receber mais colesterol livre .


A particula formada é a HDL3. A adição de material de superficie proveniente dos remanescentes dos quilomicra e VLDL e a acção da LCAT transforma-a em HDL2.

.
Transferência do colesterol
As HDL são captadas por receptores sendo os esteres do colesterol transferidos por retroendocitose ou por translocação.


cortesia de S.Conova
Na translocação as HDL nativas são captadas pelos receptores dos macrófagos e recebem o colesterol, tomando uma forma esférica

A retroendocitose é uma endocitose mediada por receptor. As HDL são captadas pelos macrófagos e recebem os esteres do colesterol. São necessários a CETP e a apo CI .

Intercambio das apoproteinas
Como vimos, no decorrer do metabolismo há um intercambio constante de apoproteinas.
As apo B transportam o colesterol para os tecidos periféricos enquanto as apo E promovem o retorno do colesterol para o fígado.
As HDL são um depósito de apo C que quando cedida aos quilomicra e VLDL activa a lipoproteina lipase.
Pelo contrário as HDL recebe apo AI das partículas residuais que irá activar a LCAT.
Lipoproteinas e ateroesclerose

Capitulo 4
RECEPTORES
RECEPTORES
O conceito de receptores para as lipoproteinas deve-se a GOLDSTEIN e BROWN que demonstraram que a maioria das LDL só se catabolisava após fixação a receptores existentes na membrana celular.
Posteriormente constatou-se que para lá destes receptores que reconhecem as apoproteinas B e E ( receptores BE) há outros tipos de receptores.
Receptores BE
Estrutura
É uma glicoproteina com 893 aminoácidos distribuída por cinco domínios
Zonas funcionais
O receptor tem duas zonas funcionais, uma de ligação situada no folheto externo da membrana e uma de internalização no interno
Devido à constituição em aminoácidos da zona de internalização e à afinidade de uma proteína membranária, a clatrina, os receptores acumulam-se em poços revestidos (coated pits)
Síntese dos receptores
Nos ribossomas sintetiza-se um precursor dos receptores que migra para o aparelho de Golgi.
No aparelho de Golgi sofre alterações pós-translaccionais como a glicosilação.
O gene encontra-se no cromossoma 19..
Coated pits e coated vesicles
Quarenta e cinco minutos após a sua síntese o receptor aparece na membrana, agregando-se em vesículas revestidas por distribuição ao acaso.
As coated pits interactuam com as LDL formando-se dentro de 3-5 minutos vesículas de endocitose revestidas de clatrina

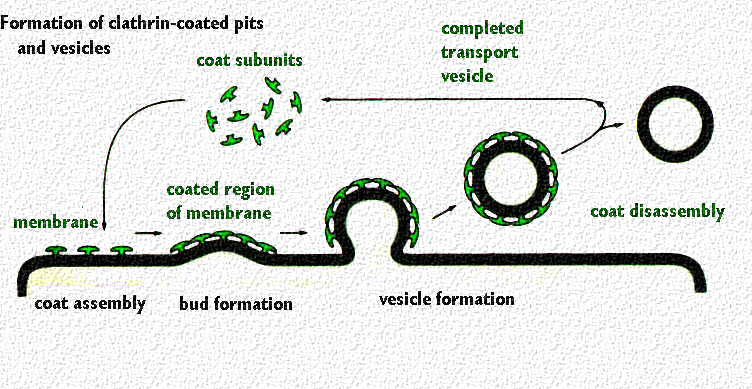
cortesia de Gwen Childs
Univ. of Arkansas for Medical Sciences
Estas vesículas perdem rapidamente a clatrina e fundem-se com outras para formar vesículas maiores de contorno irregular – endossomas e receptossomas

cortesia de Gwen Childs
Univ. of Arkansas for Medical Sciences
Reciclagem dos receptores
As LDL separam-se dos receptores a um pH inferior a 6,5, voltando estes para a superfície


cortesia de Gwen Childs
Univ. of Arkansas for Medical Sciences
Lisossomas
As LDL são captadas pelos lisossomas.
As proteínas são hidrolisadas em aminoácidos e o colesterol esterificado em colesterol e ácidos gordos


Cortesia de
Gwen V.Childs
Univ. of Arkansas for Medical Sciences
Sinopse

Regulação
A captação de colesterol pelos receptores tem por objectivo fornecer à membrana celular o colesterol que ela necessita para a sua estabilidade.
A captação do colesterol está regulada para que seja fornecida à membrana uma quantidade necessária mas não excessiva.
Os mecanismos reguladores exercem-se sobre:
HMG CoA redutase
· Repressão da sua síntese
· Aceleração do seu catabolismo
· Inibição alosterica
ACAT
Aumento da sua actividade ficando os esteres do colesterol depositados no citoplasma como gotículas de gordura.
Receptores
Repressão da sua síntese.
Receptores BE doutros tecidos
Os receptores BE encontram-se não só nos fibroblastos mas em muitos órgãos e tecidos.
A actividade especifica mais elevada foi encontrada no córtex suprarenal e corpo amarelo devido ao colesterol ser precursor das hormonas esteroides.
Devido à sua massa, o fígado é o maior possuidor de receptores.
As células endoteliais tem receptores que não internalizam as LDL, a não ser que estejam lesados.
Os macrofagos não têm receptores para as LDL mas captam LDL modificadas.
Lipoprotein Related Protein (LPR)
Tem cinco domínios funcionais com alguma semelhança ao BE.
Não reconhece a apo B.
Liga-se fracamente às VLDL.
Têm alta finalidade para os remanescentes das VLDL, ricos em apo E.
Receptores para as VLDL
O seu papel não é ainda bem conhecido.
Atribui-se-lhes um papel importante na captação de remanescentes ricos em apo E.
Podem mediar a captação de outros ligandos como a proteína de ligação da riboflavia, a a2 macroglobulina e a lactoferrina.
Receptores 2 das apo E
Fazem a captação das partículas ricas em apo E nos astrociros
Receptores das HDL
Medeiam a endocitose e a translocação
Capitulo 4
DISLIPEMIAS
DISLIPEMIAS
Bibliografia geral
Classificação de Frederickson-OMS
É uma classificação baseada na ordem de migração electroforetica:
· Tipo I – quilomicra
· Tipo IIa – beta-lipoproteinas
· Tipo III – IDL, ocupando uma zona que faz coalescer as b e as pré-b
· Tipo IV – pré-beta
· Tipo V – coexistência de pré-beta e quilomicra
· Tipo IIa - coexistência de beta e pré-beta
Classificação de Frederickson


Tipo I
Há aumento dos quilomicra e portanto hipertrigliceridémia.
Um processo simples e eficaz de reconhecer os quilomicra consiste no exame do soro após 24 horas de repouso a 4 graus.
Os quilomicra aparecem como um sobrenadante cremoso num soro límpido.
Este tipo é muito raro, não tendo sido descrito nenhum caso em Portugal.
Deve-se à falta da lipoproteina lipase ou da carência da apo CII, activadora da lipoproteina lipase

Tipo IIa
Há aumento das LDL com a consequente hipercolesterolémia
Há uma forma dependente da alimentação ( hipercolesterolémia poligénica) e outra devida à diminuição ou ausência de receptores
Tipo IV
Aumentam apenas as VLDL, havendo portanto uma hipertrigliceridémia.
O soro encontra-se turvo.
Pode ser devido a:
- · Resistência periférica à insulina com um hiperinsulinismo secundário com um aumento de lipólise e portanto maior produção de triglicéridos

- Diminuição da destruição de triglicéridos por carência em apo C

Tipo III
Há acumulação de IDL que se manifesta na electroforese por uma banda larga( broad beta) que engloba as bandas das beta e pré-beta.
O colesterol e triglicéridos estão elevados.


Tipo V
É a associação dos tipos I e IV, ou seja, aumento dos quilomicra e VLDL com a consequente hipertrigliceridémia.
O soro apresenta um sobrenadante cremoso( quilomicra) e um infranadante turvo (VLDL).
Tipo IIb
É a associação dos tipos II e IV, implicando assim o aumento de colesterol e triglicéridos.
Criticas a esta classificação
Na ideia de Frederickson esta classificação referia-se apenas a fenotipos de uma alteração do genotipo – cada tipo seria a expressão de uma mutação, ou seja de um genotipo
Actualmente este conceito não pode ser aceite pelas seguintes razões:
· Nem sempre a árvore genealógica mostra tratar-se do mesmo fenotipo

· Após o tratamento é frequente observar mudança dos tipos, particularmente Iib em IV.
· Há dislipémias, as dislipémias normolipémicas que não entram nesta classificação.
Classificação de De Gennes
Esta classificação segue critérios bioquímicos e precisa alguns critérios:
Associação de varias dislipemias
Pode haver num mesmo doente associação de varias dislipemias
Hipercolesterolemias
Bibliografia
Directorios
http://www.emedicinehealth.com/high_cholesterol/article_em.htm
Textos
LDL
Hipercolesterolemia poligenica
Há uma associação entre excesso de lípidos alimentares, nomeadamente ácidos gordos saturados e uma alteração genética da homeostase do colesterol.
O colesterol é inferior a 300.
É sensível à dieta.
Hipercolesterolemia familiar
São hipercolesterolémias elevadas de origem genética, pouco sensíveis à dieta.
Nas formas heterozigoticas, o colesterol situa-se entre 300 e 400 e nas homozigoticas acima de 400.
São quase sempre devidas a alterações dos receptores BE.
Acompanham-se de xantomas cutâneos e tendinosos.
Os homozigoticos desenvolvem doença coronária antes dos 30 anos, sendo o prognostico grave.
As formas heterozigoticas tratam-se com estatinas.
As homozigoticas podem ser tratadas com sequestrantes dos ácidos biliares e colesterol-aferese.
Bibliografia
Hipertrigliceridémias
As VLDL e os triglicéridos estão aumentados, mas não os quilomicra.
O soro encontra-se turvo.
Hipertrigliceridémias primarias
Hipertrigliceridémia familiar
Triglicéridos aumentados na ausência de outras causas
Existe noutros membros da família
É devida à hiperprodução de VLDL
Sindroma de deficiência em HDL
Hipertrigliceridémia moderada
Grande baixa das HDL
Hipertensão dislipémica familiar
Hipertensão antes dos 60 anos
Hipertrigliceridémia
Ocorre em 15% dos hipertensos
Hipertrigliceridemia com remanescentes
Remanescentes semelhantes às IDL
Bibliografia
Hipertrigliceridémias secundárias
Podem-se observar na obesidade, alcoolismo, diabetes e insuficiência renal
Hiperlipemia mista
Triglicéridos e colesterol aumentados.
Embora as alterações lipídicas muitas vezes não sejam grandes, acarreta um risco aumentado, especialmente na diabetes tipo 1.
Quilomicronemias
Correspondem ao tipo I de Frederickson.
Há aumento isolado de quilomicra.
Num soro mantido 24 horas no frigorifico, os quilomicra sobrenadam.
Devem-se a uma deficiência da liproteina lipase.
São muito raras.
Dislipemias familiares com IDL( tipo III)
Há uma acumulação de IDL reconhecível na electroforese por uma banda englobando as beta e pré-beta (broad beta) e pelas suas caracteristocas na ultracentrifugação(floating beta).
O colesterol e trigliceridos estão aumentados.
Deve-se a uma homozigotia do alelo E2, que tem uma capacidade deficiente de ligação aos receptores, ao contrario dos E3 e E4.
Dislipemias normolipemicas
Tipo IV latente
Designação introduzida por HALPERN em 1973 para designar aumento da pré-beta com trigliceridos normais.
Os triglicéridos surgem aumentados quando doseados três horas após uma refeição
As VLDL têm uma composição alterada
Apoproteinas
Há casos de aumentos isolados de apo B.
Lp(a)
É uma variante das LDL, sendo a sua síntese controlada geneticamente.
Bibliografia
Isoformas da apo E
Descreveram-se isoformas da apo E com normolipémia.
Dislipemias secundarias
Diabetes
Diabetes e mortalidade cardiovascular
A mortalidade cardiovascular representa 70 a 75% da mortalidade dos diabéticos, sendo maior nas mulheres
O papel da insulina no metabolismo das lipoproteinas é independente dos seus efeitos sobre a glicose.
Modula a lipoproteina lipase e pode influenciar a LCAT.
Os trigliceridos estão aumentados nas diabetes.
O colesterol está nornal.
As HDL estão significativamente mais baixas.
As apo AI estão baixas no tipo I.
As apo CII e CIII estão aumentadas.
Nas mulheres as alterações lipidicas são maiores e nunca recuperam totalmente.
A dislipemia representa um factor de risco importante pois que a mortalidade cardiovascular representa 70% das causas de mortalidade da diabetes.
Suprarenal
Os corticoides provocam uma resposta bifásica nos lipidos – descida seguida de subida
No Cushing e em doentes tratados com corticoides os triglicéridos estão elevados e associados a uma maior captação de ácidos gordos livres
Tiroideia
No hipotiroidismo há hipercolesterolémia e por vezes hipertrigliceridemia por uma captação deficiente de LDL pelos receptores e actividade diminuída da lipoproteina lipase.
Colestase
Há uma hiperlipemia marcada causada por uma lipoproteina anormal, a lipoproteina X.
Há uma hiperlipemia marcada causada por uma lipoproteina anormal, a lipoproteina X.
Há uma hiperlipemia marcada causada por uma lipoproteina anormal, a lipoproteina X.
Nefropatias
A insuficiência renal crónica pode estar associada com hipertrigliceridémia e hiperquilomicronemia.
A lipoproteina lipase tem uma actividade diminuída devido à existência de um seu inibidor.
Os trigliceridos estão associados com a clearance de creatinina.
A hemodiálise não melhora esta situação.
As HDL estão baixas, sendo a sua subida após uma transplantação um sinal de bom prognóstico.
Na nefrose há hipercolesterolémia.
Na hemodiálise há hipertrigliceridemoa.
Capitulo 5
HIPOLIPEMIAS
Deficit em apo B
Noções gerais
As apo B são necessárias para a síntese dos quilomicra.
São transportadoras de vitaminas lipossoluveis especialmente as D e A.
É de considerar que os ácidos gordos de media cadeia são absorvivos directamente pelo fígado, tendo outra metaboização. A sua administração será portanto útil nesta situação.
Será também útil administrar suplementos de vitaminas A e D.
A-betalipoproteinemia
É uma forma autossomica recessiva.
Faltam as Apo B-100 e B-48.
Há ausência de quilomicra, VLDL e LDL.
Hipobetalipoproteinemia
É uma forma autossomica dominante
Forma homozigotica
Ausência de VLDL
Diminuição das LDL e HDL
Forma heterozigotica
Diminuição de 50% das LDL e VLDL
Forma heterozigotica
Diminuição de 50% das LDL e VLDL
A-betalipoproteinemia normotrigliceridemica
Falta a apo- 100 mas não a apo B-48
Ausência de LDL e VLDL
Quilomicra em quantidades normais
Deficiência em Apo A-I
Hipo-alfalipoproteinemia familiar
Autosomica dominante.
Teores de HDL e Apo AI 50% do normal.
Predisposição para a ateroesclerose prematura.
Doença de Tangier
Ausência quase total de HDL.
Anomalia do gene da Apo A-I que impede a convrsão da pré-pro-apo A-I.
Bibliografia
Deficiência familiar em apo E
Deficiência em apo C-II
Doença autossomica recessiva.
Defiiciencia total ou grande de apo C-II.
Sem comentários:
Enviar um comentário