Via final
A neoglicogénese não é o inverso da glicólise porque nem todas as reacções reversíveis devido a barreiras energéticas.
Os enzimas que funcionam numa só direcção são os enzimas chave( glicolíticos ou glicogénicos).
Os enzimas que funcionam nas duas direcções são os enzimas bifuncionais.
Enzimas glicoliticos chave
Barreiras energéticas impedem que algumas reacções sejam reversíveis .São as reacções catalisadas por
- Hexocinases
- Fosfofrutocinase 1
- Piruvato cinase
Enzimas glicogenico chave
São enzimas que catalisam as reacções inversas dos enzimas glicogenicos chave:
¨ Formação do ácido enolpirúvico
¨ Desforilação da fructose –1,6- bisfosfato em frutose-6-fosfato
¨ Passagem de glicose-6- fosfato a glicose
Formação do ácido fosfoenolpirúvico
Esta reacção faz-se em duas étapas:
¨ Formação do ácido oxaloacético
¨ Descarboxilação fosforilante deste em ácido fosfoenolpirúvico

Formação de ácido oxaloacético
Trata-se da reacção de Wood e Werkman, catalisada pela piruvato carboxilase.
Necessita de ATP.
Tem a biotina como coenzima.
A fixação da biotina ao enzima é activada pelo acetil-CoA
CO2
ATP
Acido pirúvico --------------- Acido oxaloacetico
Piruvico carboxilase
Descarboxilação fosforilante do ácido oxaloacético
O fosfato de alto potencial necessário para esta reacção vem do GTP
.O enzima que catalisa esta reacção é a fosfoenolpiruvato carboxicinase
Travessia das mitocondrias
O ácido oxaloacético forma-se nas mitocôndrias mas a sua transformação em fosfoenolpirúvico é extramitocondrial.
Como o oxaloacético não atravessa a membrana mitocondrial, terá que se transformar num composto que atravesse a membrana e em seguida se reconverta nele.
Há duas vias – a via do malato e a via do aspartato.
Via do malato
O oxaloacetato é reduzido em malato pela malatodeidrogenase mitocondrial na presença de NADH.
A translocase dos ácidos dicarboxílicos faz o malato abandonar as mitocôndrias para este no citoplasma dar de novo oxaloacetato.
Via do aspartato
A aspartatoaminotransferase transamina o oxaloacético em ácido aspártico.
Via final
A neoglicogénese não é o inverso da glicólise porque nem todas as reacções reversíveis devido a barreiras energéticas.
Os enzimas que funcionam numa só direcção são os enzimas chave( glicolíticos ou glicogénicos).
Os enzimas que funcionam nas duas direcções são os enzimas bifuncionais.
Enzimas glicoliticos chave
Barreiras energéticas impedem que algumas reacções sejam reversíveis .São as reacções catalisadas por
- Hexocinases
- Fosfofrutocinase 1
- Piruvato cinase
Enzimas glicogenico chave
São enzimas que catalisam as reacções inversas dos enzimas glicogenicos chave:
¨ Formação do ácido enolpirúvico
¨ Desforilação da fructose –1,6- bisfosfato em frutose-6-fosfato
¨ Passagem de glicose-6- fosfato a glicose
Formação do ácido fosfoenolpirúvico
Esta reacção faz-se em duas étapas:
¨ Formação do ácido oxaloacético
¨ Descarboxilação fosforilante deste em ácido fosfoenolpirúvico
Formação de ácido oxaloacético
Trata-se da reacção de Wood e Werkman, catalisada pela piruvato carboxilase.
Necessita de ATP.
Tem a biotina como coenzima.
A fixação da biotina ao enzima é activada pelo acetil-CoA
CO2
ATP
Acido pirúvico --------------- Acido oxaloacetico
Piruvico carboxilase
Descarboxilação fosforilante do ácido oxaloacético
O fosfato de alto potencial necessário para esta reacção vem do GTP
.O enzima que catalisa esta reacção é a fosfoenolpiruvato carboxicinase

cortesia de Joyce Diwan
Travessia das mitocondrias
O ácido oxaloacético forma-se nas mitocôndrias mas a sua transformação em fosfoenolpirúvico é extramitocondrial.
Como o oxaloacético não atravessa a membrana mitocondrial, terá que se transformar num composto que atravesse a membrana e em seguida se reconverta nele.
Há duas vias – a via do malato e a via do aspartato.
Via do malato
O oxaloacetato é reduzido em malato pela malatodeidrogenase mitocondrial na presença de NADH.
A translocase dos ácidos dicarboxílicos faz o malato abandonar as mitocôndrias para este no citoplasma dar de novo oxaloacetato.
Via do aspartato
A aspartatoaminotransferase transamina o oxaloacético em ácido aspártico
.
Via final
A neoglicogénese não é o inverso da glicólise porque nem todas as reacções reversíveis devido a barreiras energéticas.
Os enzimas que funcionam numa só direcção são os enzimas chave( glicolíticos ou glicogénicos).
Os enzimas que funcionam nas duas direcções são os enzimas bifuncionais.
Enzimas glicoliticos chave
Barreiras energéticas impedem que algumas reacções sejam reversíveis .São as reacções catalisadas por
- Hexocinases
- Fosfofrutocinase 1
- Piruvato cinase
Enzimas glicogenico chave
São enzimas que catalisam as reacções inversas dos enzimas glicogenicos chave:
¨ Formação do ácido enolpirúvico
¨ Desforilação da fructose –1,6- bisfosfato em frutose-6-fosfato
¨ Passagem de glicose-6- fosfato a glicose
Formação do ácido fosfoenolpirúvico
Esta reacção faz-se em duas étapas:
¨ Formação do ácido oxaloacético
¨ Descarboxilação fosforilante deste em ácido fosfoenolpirúvico
Formação de ácido oxaloacético
Trata-se da reacção de Wood e Werkman, catalisada pela piruvato carboxilase.
Necessita de ATP.
Tem a biotina como coenzima.
A fixação da biotina ao enzima é activada pelo acetil-CoA
CO2
ATP
Acido pirúvico --------------- Acido oxaloacetico
Piruvico carboxilase
Descarboxilação fosforilante do ácido oxaloacético
O fosfato de alto potencial necessário para esta reacção vem do GTP
.O enzima que catalisa esta reacção é a fosfoenolpiruvato carboxicinase
cortesia de Joyce Diwan
Travessia das mitocondrias
O ácido oxaloacético forma-se nas mitocôndrias mas a sua transformação em fosfoenolpirúvico é extramitocondrial.
Como o oxaloacético não atravessa a membrana mitocondrial, terá que se transformar num composto que atravesse a membrana e em seguida se reconverta nele.
Há duas vias – a via do malato e a via do aspartato.
Via do malato
O oxaloacetato é reduzido em malato pela malatodeidrogenase mitocondrial na presença de NADH.
A translocase dos ácidos dicarboxílicos faz o malato abandonar as mitocôndrias para este no citoplasma dar de novo oxaloacetato.
Via do aspartato
A aspartatoaminotransferase transamina o oxaloacético em
ácido aspártico.
Uma translocase transfere o
ácio aspártico para o citoplasma
onde se converte de novo em oxaloacético

Desfosforilação da frutose-1,6-bisfosfato
A frutose-1,6-bisfosfatase catalisa esta reacção.
O ADP é um efector alostérico negativo ao mesmo tempo que
activa a fosfofrutocinase.
O enzima é activado pelo ácido láctico e pelo cortisol.
Desfosforilação da glicose-6-fosfato
A hidrólise irreversível do fosfato é catalisada pela
glicose-6-fosfatase.
O enzima está fortemente ligado à membrana
A glicose-6-fosfato é transportada para o retículo por um transportador, sendo
aí hidrolisada
A glicose formada é transportada de novo para o citossol por
um transportador
Necessita de
fosfolípidos para a sua actividade.
É inibido pelos fosfatos e pela glicose
Necessita de três transportadores:
Necessita de três transportadores
1. Transporte
d a glicose-6-fosfato para o lume
2. Transporte
do fosfato para o citossol
3. Transporte
da glicose para o citossol
Ciclo dos Cori
O músculo não consegue
transformar em piruvato o ácido láctico formado após um esforço muscular
intenso.
Para se converter em piruvato, o ácido láctico tem que
ser transportado para o fígado ou rim,
que têm enzimas que permitem fazer esta conversão- é o ciclo dos Cori

Ciclo de Fehlig
Uma outra forma de
metabolização do acido piruvico
pelo musculo para fins glicogenicos é a sai conversão em alanina
A alanina
dirigir-se-á para o fígado, onde será reconvertida em acido piruvico
É o ciclo de
Fehlig


Regulação
A glicolise e a neoglicogenese são controladas pelos mesmos
mecanismos, para que funcione apenas uma das vias
A fosfofrutocinase 1 é
estimulada pelo AMP e inibida pelo ATP e citratos, efectores que têm uma acção
oposta sobre a frutose-1,6-bisfosfatase
Assim, quando há um baixo nível energético, a glicolise é
estimulada e no caso contrario é estimulada a gliconeogenese
A frutose-1,6-
bisfosfato tem níveis baixos na inanição e elevados na saciedade
O antagonismo glucagina-insulina implica a estimulação da
gliconeogenese
Assim, quando há um baixo nível energético, a glicolise é
estimulada e no caso contrario é estimulada a gliconeogenese
A piruvatocinase
é inibida pela ATP e Alina, ao contrario da carboxicinase
Capitulo 12
REGULAÇÃO DO METABOLISMO DO GLICOGENIO
A regulação do metabolismo do glicogénio faz-se através de
dois enzimas fundamentais, a glicogénio sintase e a fosforilase.
O AMP cíclico desempenha um papel fundamental na regulação
destes enzimas pois mediante a fosforilação destes enzimas inibe a sintase e
estimula a fosforilase actuando assim no
sentido da glicogenólise.
AMP cíclico
O AMP cíclico actua no metabolismo do glicogénio no sentido
da glicogenolise pois fosforila a forma a da glicogénio sintetase, tornando-a
inactiva e activa a forma a da fosforilase

RESUMINDO:
GLUCAGINA ( fosforilação)
Fosfatase a
Sintase b
GLICOLISE
INSULINA ( desfosforilação)
Fosfatase b
Sintase a
GLICOGENESE

Inibidor proteico I
A glicogenólise é uma forma rápida de mobilização da glicose
que só deverá funcionar quando for necessário mobilizà-la ràpidamente.
Esta regulação faz-se pelo balanço
fosforilação-desfosforilação, encaminhando a fosforilação para a glicogenólise
e a desfosforilação para a glicogénese.
A desfosforilação é assegurada pela fosfoproteina fosfatase
I que desfosforila a glicogénio sintase, a fosforilase e a cinase.
Este enzima é inibido pelo inibidor proteico I que se
forma por fosforilação da sua forma inactiva pela acção da proteina cinase
formada pelo AMP cíclico.
O inibidor é inactivado ao ser desfosforilado por uma
fosfoproteina fosfatase I.
RESUMINDO: Ao ser activado o inibidor proteico I activa
todos os enzimas que levam à glicogenólise passando-se o contrário quando é
desfosforilado.
Capitulo 13
PATOLOGIA DA NEOGLICOGENESE
Deficiência em Glicose-6-fosfatase
É a glicogenose tipo I
Deficiência em
frutose-1,6-bisfosfatase
Sintomas
Hiperventilação
Convulsões
Coma
Hiperventilação
Convulsões
Coma
Tratamento
Tratamento das crises
agudas pela infusão intravenosa de glicose
Evitar jejum
Restrição de frutose e sacarose
Para prevenção da hipoglicemia dar um glucido de libertação
lenta como o amido de milho
Deficiência em
piruvico carboxilase
Consequência da
deficiencia
Deplecção de oxaloacetato
acarretando níveis reduzidos de aspartato, metabolito necessário para a síntese
da ureia
Sintomas
Acidose láctica
Hiperamoniemia
Tratamento
Suplementos de aspartato e citrato
Capitulo 14
Regulação da glicemia
O fígado dos mamíferos é capaz de responder a níveis
diferentes de glicose circulante. Quando o teor em glicose do sangue portal é
alto, o fígado absorve mais glicose. Quando é baixo, liberta glicose.
Como as células hepaticas são totalmente permeáveis à
glicose, pensa-se que o balanço entre a absorção e a libertação de glicose se
deve à actividade dos enzimas glicoliticos e glicogenicos chave.
Na regulação da glicemia as hormonas desempenham um papel
importante.
AMP cíclico
Muitas hormonas actuam sobre o AMP cíclico
Insulina
A insulina inibe a formação do AMP cíclico
Combina-se com um receptor que induz a síntese de um segundo
mensageiro que inactiva a cinase
Induz os enzimas glicolitico chave inibe os glicogenico chave
Induz os enzimas da
lipogenese e inibe os lipoliticos ( acção sobre o AMP cíclico)
Alem disso facilita a entrada da glicose na celula
Favorece a
glicogenese



Adrenalina
A adrenalina estimula a produção de AMP cíclico
combinando-se com uma proteina específica existente no interior da membrana, o receptor
adrenérgico.
O complexo receptor- adrenalina na presença de ATP
combina-se com a proteina G estimulando
a adenilciclase.
A adrenalina actua apenas no músculo e não no fígado

A adrenalina actua apenas no músculo e não no fígado
Glucagina
No fígado a glucagina toma o lugar da adrenalina.
Como resposta a uma descida da glicose sanguínea as células
do pâncreas segregam glucagina que combinando-se a um receptor estimula a
adenilciclase por um mecanismo semelhantre ao da adrenalina, favorecendo assim
a glicogenólise

Na figura seguinte estão esquematizadas as acções da
insulina e glucagina na regulação da glicemia

Glicocorticoides
- Activam
o catabolismo das proteínas e lipidos, favorecendo a neoglicogenese
- O
acetil CoA formadono catabolismo
dos lipidos activa a piruvato carboxilase, enzima glicogenico chave, e
inibe os glicoliticos chave
- Induzem a síntese dos enzimas glicogenico chave
Capitulo 15
DIABETES
Conceitos
A diabetes mellitus é uma doença heterogenica caracterizada
pela presença de hiperglicemia
A hiperglicemia é
devida a uma deficiência da acção da insulina
As duas principais causas são a menor produção de insulina
pelo pâncreas ou uma resposta deficiente
da insulina nos órgãos alvo
Estas duas causas definem dois tipos de diabetes – tipo 1 e
tipo 2 respectivamente
Diabetes tipo 1
Causas
É causada pela destruição autoimune das células beta dos
ilhéus de Langerhans
A reacção inicia-se por um mecanismo desconhecido
A destruição das células beta desencadeia uma menor produção
de insulina com hiperglicemia e os outros sinais de diabetes
Parece ser o resultado da combinação de uma susceptibilidade
genetica com factores ambientais
Fases
Existência de autoanticorpos com glicose pós-prandial normal
Diminuição da tolerância à glicose
Hiperglicemia em jejum embora se produza ainda insulina
suficiente para produzir a cetose
A produção de insulina desce ainda mais e os doentes
tornam-se dependentes da insulina exógena
Complicações
São a aterosclerose, neuropatia periférica, insuficiência
renal, retinopatia
O desenvolvimento e gravidade destas complicações dependem
do substracto genético e do grau de controle metabólico
Um controle rigoroso da glicemia reduz o risco das
complicações de 35 a 75%
Diabetes tipo 2
Causas
Surge no adulto, quase sempres após a meia idade
Não é autoimune
A susceptibilidade genética é um requisito indispensável mas
a sua expressão clínica está determinada em parte por factores ambientais
Nos tipo 2 a resposta à ingestão de glicose é inadequada e
os padrões basais são elevados, sinais de resistência à insulina
Complicações
As mesmas do tipo 1
Tratamento
A perda de peso, o aumento do exercício físico e a supressão
do açucar são os métodos mais efectivos
Quando não resultam totalmente devem ser complementados com
hipoglicemiantes orais como as sulfanilureias ou as biguanidas
Capitulo 16
Capitulo 17
CICLO DE DICKENS-HORECKER
Conceito
Via que utiliza os seis carbonos da glicose para gerar
equivalentes redutores e pentoses
Também é conhecido
por ciclo das pentoses, via das pentose-fosfatos e desvio das hexose-fosfatos
Realiza-se no figado, tecido adiposo, córtex suprarenal, testículos, glândula mamaria
lactante e eritrocitos
Tem uma fase oxidativa e uma não oxidativa
Via oxidativa

Via oxidativa


Formação de
fosfoglicolactona
O C1 da glicose-6-fosfato perde dois hidrogénios, captados
pelo NADP
A reacção é catalisada pela glicose-6-fosfato deidrogenase
ou enzima intermediário de Warburg
É inibido por certas drogas como as sulfonamidas e a
primaquina e activada pela insulina
Mais de 100.000 pessoas têm uma deficiência hereditária neste enzima
Formação de acido fosfogluconico
A lactonase transforma a fosfoglicolactona em acido
fosfogluconico

Formação de
ribulose-5-fosfato
Forma-se uma cetona intermediaria, sendo os hidrogenios
captados pelo NADP
A cetona intermediaria é descarboxilada em
ribulose-5-fosfato

O NADPH formado é utilizado no metabolismo dos lípidos
Isomerização das
pentoses-fosfato
A fosfopentose-epimerase
transforma a ribulose-5-fosfato em xilulose-5-fosfato e a fosfopentose
isomerase isomeiza-a em ribose-5- fosfato


Transfere o C1 e C2 (cetol) da
xilulose-5- fosfato para a ribose-5-fosfato originando uma cetose, a
sedoheptulose-7-fosfato e o gliceraldeido-3-fosfato
A reacção é catalisada pela
transcetolase
Dois compostos com cinco carbonos,
originaram um com 7 e um com 3
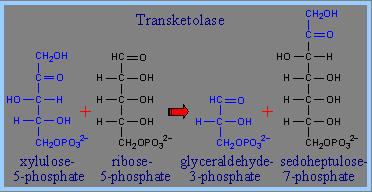
sx
Aldolização
A aldolase catalisa a
transferência de três carbonos da sedoheptulose-7-fosfato para o fosfogliceraldeido
para originar um composto com quatro carbonos (eritrose-4-fosfato) e um com
seis (frutose-6-fosfato)



(frutose-6-fosfato)

2ª transcetolização
A transcetolase efectua uma nova
recombinação os dois primeiros carbonos da xilulose para a eritrose
Forma-se frutose-6-fosfato e
fosfogliceraldeido


Equilentes redutores sob a forma de NADPH que serão utilizados em reacções de
síntese
Fornecer ribose
para a síntese dos ácidos nucleicos
Manter
a integridade da membrana dos eritrocitos pela redução do glutatião
Metabolisar
as pentoses alimentares
Regulação
O factor mais
importante é a concentração celular de
NADPH
A
disponibilidade em NADP regula a reacção limitante, a reacção da
glicose-6-fosfato-deidrogenase
PATOLOGIA DO CICLO DE DICKENS- HORECKER
Glutatião
O glutatião tem uma actividade anti-oxidante por destruír os
peróxidos
A regeneração do glutatião faz-se pela acção da glutatião
peroxidase, que necessita de NADPH
Como o ciclo de Dickens.Horecker é o principal fornecedor de
NADPH,qualquer falha deste ciclo reflectir-se-á na regeneração do glutatião e
consequente aumento do stress oxidativo
Eritrocitos
Nos eritrocitos a única fonte de NADPH é o ciclo de
Dickens-Horecker
Qualquer diminuição de produção de NADPH por este ciclo terá
sempre consequências nefastas, por falta de alternativas
As consequências serão hemolise por enfraquecimento da
parede celular e maior oxidação da hemoglobina em metahemoglobina
Deficiencia em
glicose-6-fosfato-deidrogenase
É a deficiência hereditária mais espalhada no mundo – afecta
mais de 400 milhões
Na bacia mediterrânica e em Africa está muitas vezes relacionada
com a resistência ao Plasmodio falciparum
Nas áreas em que a malária é endémica a deficiência tem uma
prevalência de 5 a 25% enquanto que em áreas não endémicas é inferior a 0,5%
A causa dos sintomas é a falta de produção de NADPH
Esta deficiência se manifesta por anemia hemolítica
Manifesta-se na presença de oxidantes
Também tem o nome de favismo por aparecer após a ingestão de
favas, que é um oxidante.
Alimentos e
medicamentos a evitar
Há alimentos e medicamentos que devem ser completamente proscritos
para impedir o aparecimento de uma nova crise
Capitulo 19
METABOLISMO DAS
HEXOSES E DA LACTOSE
A quase totalidade do metabolismo das oses e oligosidos
faz-se através da glicose
As outras hexoses podem-se converter em glicose ou em
intermediários da glicose, do mesmo modo que a glicose pode originar algumas
hexoses
Frutose
É fosforilada pela frutocinase em frutose-1-fosfato.
A frutose-1-fosfato é
cindida pela aldolase da frutose-1-fosfato em gliceraldeido e
fosfodihidroxiacetona
A triose-cinase fosforila o gliceraldeido em
fosfogliceraldeido

Galactose
Embora a galactose
seja um epimero da glicose, a sua
transformação nesta é complexa.
Primeiramente forma-se por accão da galactocinase em
galactose-1-fosfato pela acção da galactocinase
Esta transforma-se em UDP galactose pela acção da
galactose-1-fosfato uridiltransferase
A epimerização faz-se a este nível pela acção da
UDP-galactose-4-epimerase que em seguida irá originar a glicose-1-fosfato

sfato
Deficiência em
transferase
Causas
O recemnascido ingere 20% das suas calorias como lactose,
que contem galactose
Na falta deste enzima, a
galactose-1-fosfato não se metabolisa, acumulando-se nos rins, fígado e cérebro
A sua transformação em galactitol pode produzir
cataratas
Sintomas
Os recemnascidos ou lactentes
apresentam entre outros sintomas alterações hepáticas, , convulsões ou
letargia, atraso mental e cataratas
Quando o diagnostico não é feito
precocemente a cirrose, o atraso mental e as cataratas tornam-se irreversiveis
Cataratas
É um dos sintomas mais frequentes
Deve-se à conversão da galactose no açúcar álcool galacitol
por uma galactose redutase dependente do NADPH
Esta redutase só existe no tecido nervoso e no cristalino
A níveis circulantes normais de galactose, a actividade do
enzima não causa efeitos lesivos.
A concentrações elevadas o galacitol cria tumefacção
osmotica do cristalino com a consequente formação de cataratas
Tratamento
O despiste desta doença permite o
tratamento precoce
A eliminação precoce da galactose
cura os sintomas
Deficiência em
galactocinase
Os principais metabolitos acvumulados são a galactose e o
galactitol
O unico sintoma é representado pelas cataratas
O tratamento é restrição da galactose
Deficiência da
UDPGalactose epimerase
Os sintomas são
semelhantes aos da deficiência em transferase
Está indicada a restrição da galactose
Frutose
Frutosuria essencial
Falta a fosfofrutocinase hepática
A situação é benigna porque não se acumulam produtos tóxicos
Parte da frutose ingerida
é fosforilada parcialmente pela
hexocinase, entrando na glicolise
Intolerância
hereditária à frutose
Causas
Falta a aldolase
Acumula-se frutose-1-fosfato em vários tecidos, inibindo a
glicogeneolise e a gliconeogenese
A frutose necessita de ATP para ser fosforilada.
A acumulação de
frutose-1-fosfato leva portanto a uma deplecção de ATP
A falta de ATP impede o fígado de realizar as suas funções
normais
principalmente por
impedir o funcionamento das bombas dependentes de ATP
Sintomas
Os sintomas só surgem após a administração de frutose ou
sacarose
A ingestão aguda de
frutose produz hipoglicemia
A ingestão crónica produz atraso de crescimento e
insuficiência hepática
Se a ingestão de frutose se mantiver surgem crises hipoglicemicas recidivantes e
insuficiências hepática e renal que podem levar à morte
Tratamento
Eliminação total da frutose, sacarose e sorbitol
Capitulo 21
METABOLISMO DOACIDO GLICURONICO
Síntese
A glicose-1-fosfato combina-se com a UTP para dar a UDPG
A UDPG oxida-se no C6 para dar o UDP-glicuronato pela
acção da UDPG deidrogenase
Esta, perdendo a UDP origina o acido glicuronico
Catabolismo
Transforma-se em L -xilulose com a formação intermediaria dos
ácidos gulonico e cetogulonico
A L-Xilulose isomeriza-se em D-xilulose que entrará no ciclo
de Dickens-Horecker

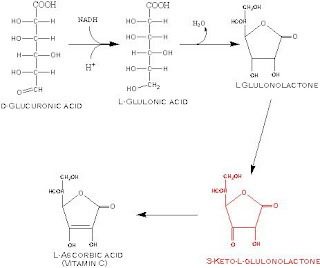
Síntese do acido ascórbico
Os primatas perderam a capacidadede sintetizar o acido
ascórbico
Os animais que o sintetizam fazem-no a partir do acido
gulonico
PATOLOGIA DOS ACIDOS URONICOS
Pentosuria idiopatica
Actividade reduzida da L-xilulose reductase
Grande excreção de pentoses pela urina, especialmente após
ingestão de acido glicuronico
É assintomática
Icterícia do
recemnascido
<
<
¨
<
Isoprenos e terpenos
Isopreno
Estrutura
O isopreno é o metilbutadieno, ou seja, um hidrocarboneto não saturado ramificado com cinco carbonos

Isopreno e sínteses biologicas

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Muitos compostos orgânicos têm um número de carbonos múltiplos de cinco.
Em 1879, BOUCHARDAT polimerizou o isopreno, formando borracha sintética.
RUCICKA propôs que muitos compostos biológicos poderiam provir do isopreno.
Esta hipótese está hoje provada, tendo-se provado que a forma activa do isopreno é o isopentilpirofosfato

{kind=link}
http://en.wikipedia.org/wiki/Terpene
Derivados preterpenicos
Resultam da condensação de um isopreno com um composto de natureza diferente.
Um exemplo é o ácido lisérgico, condensação do isopreno com a triptamina
A LSD é a dietilamida do ácido lisérgico.

http://pt.wikipedia.org/wiki/LSD
Terpenos
Estrutura
São múltiplos do isopreno
Classificam-se conforme o número de unidades
Terpenos
Terpenes
|
Isoprene
units |
Carbon
atoms | |
1
|
Monoterpenes
|
2
|
10
|
2
|
Sesquiterpenes
|
3
|
15
|
3
|
Diterpenes
|
4
|
20
|
4
|
Sesterpenes
|
5
|
25
|
5
|
Triterpenes
|
6
|
30
|
6
|
Carotenoids
|
8
|
40
|
7
|
Rubber
|
> 100
|
> 500
|
<
Sem comentários:
Enviar um comentário