METABOLISMO DOS LIPIDOS
Capitulo 1
CATABOLISMO DOS ACIDOS GORDOS
Beta-oxidação
Os ácidos gordos têm que se transformar na sua forma activa,
o acil-CoA
É a única reacção que necessita de ATP
A reacção é
catalisada pela acetil-CoA sintetase ou tiocinase
Faz-se no retículo
para os ácidos de longa cadeia e nas mitocondrias para os de curta ou media
cadeia
Entrada nas
mitocondrias
Os acil-CoA não atravessam a membrana mitocondrial, sendo
transportados pela carnitina

A acilcarnitina é transferida para a superficie interior,
onde a CPT II separa o acilo da carnitina
Etapas
1ª desidrogenação
Forma-se um acil-CoA insaturado pela acção da acil-CoA
deidrogenase
Õ enzima é uma flavoproteina
O hidrogénio é transportado pela flavoproteina transportadora de electrões (EFT) para a
ubiquinona
http://library.med.utah.edu/NetBiochem/FattyAcids/5_2a.html





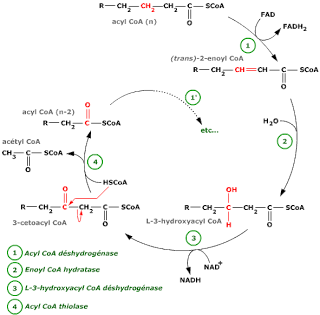


Hidratação
O acil-CoA
insaturado é hidratado em beta-hidroxiacilCoA
O enzima é a enoil-CoA hidratase
ou crotonase
.
2ª desidrogenação
O carbono beta é desidrogenado pela beta-hidroxiacil-CoA
deidrogenase formando-se o beta-cetoacil-CoA
O enzima é especifico para
o isómero L
Os hidrogénios são captados pelo NAD
Cisão
A tiolase cinde o cetoacil num acetil-CoA com menos dois
carbonos
Repetição do ciclo.
O acil-CoA sofre de
novo as mesmas etapas, formando-se de cada vez um acil-CoA com menos dois
carbonos

Se representarmos por x o numero de pares de carbonos haverá x-1 ciclos

cortesia de R.Harris

Balanço energético


cortesia de E.Harris
Regulação
Disponibilidade de
ácidos gordos
Os ácidos gordos resultam essencialmente da hidrolise dos lipidos por lipases
Actividade oxidativa
da célula
A beta-oxidação aumenta quando é necessária energia (ATP
baixo) e baixa no caso contrario
Capacidade do ciclo
de Krebs em metabolizar o acetil-CoA
Quando tal não
acontece formam-se corpos cetonicos
Beta-oxidação nos peroxissomas
Faz-se em ácidos gordos com 10 a 22 carbonos,
particularmente o acido erucico( C22:1)
Como os peroxissomas não contêm acil-CoA, os ácidos gordos
atravessam os peroxissomas por transporte activo
O acido gordo é acilado
pela acil-CoA sintetase e depois oxidado em transenoil-acil-CoA por uma
acil-CoA oxidase
Não se forma ATP porque a flavoproteina reduzida formada nesta reacção é oxidada directamente pelo oxigénio molecular com formação de agua oxigenada
Não se forma ATP porque a flavoproteina reduzida formada nesta reacção é oxidada directamente pelo oxigénio molecular com formação de agua oxigenada
Um enzima bifuncional com acções de enoil-CoA hidratase e hidroxi-acildeidrogenase forma
sucessivamente o hidroxiacil-CoA e o cetoacil-CoA
O cetoacil-CoA é
cindido pela tiolase
Inicia-se um novo ciclo que irá acabar no octanoil-CoA
O octanoil-CoA difunde-se para o citoplasma como
octanoil-carnitina, continuando aí a beta-oxidação

Alfa e ómega – oxidações
Alfa-oxidação
Dà-se nos acido-alcoois
Originam um acido gordo com menos um carbono
beta-oxidação

Doença de Refsum
Nesta doença não se faz a alfa oxidaçãovdo acido fitanico da clorofila
Ómega-oxidação
Faz-se no carbono
ómega
O enzima é uma oxigenase de função mista necesitando de NADPH e citocromo P-450
Faz-se nos microssomas
Não liberta energia



Reaction
type
|
Enzyme
|
Description
|
Reaction
|
The
first step introduces a hydroxyl group onto the ω carbon. The oxygen for the group comes
from molecular oxygen in a complex reaction (CYP-4A) that involves cytochrome P450 and the electron donor NADPH.
|
 |
||
 |
|||
The
third step is the oxidation of the aldehyde group to a carboxylic acid by NAD+. The product of
this step is a fatty acid with a carboxyl group at each end.
|
 |
After the three steps, either end of fatty acid can be
attached to coenzyme A. The
molecule can enter the mitochondrion and undergo β oxidation. The final products after successive
oxidation include succinic acid, which
can enter citric acid cycle,
and adipic acid.
Ácidos com um numero impar de carbonos
Na final da beta-oxidação forma-se um acil-CoA com três
carbonos, o propionil-CoA
O propionil-CoA transformar-se-á em succinil –CoA
Reaction
type
|
Enzyme
|
Description
|
Reaction
|
The
first step introduces a hydroxyl group onto the ω carbon. The oxygen for the group comes
from molecular oxygen in a complex reaction (CYP-4A) that involves cytochrome P450 and the electron donor NADPH.
|
|
||
|
|||
The
third step is the oxidation of the aldehyde group to a carboxylic acid by NAD+. The product of
this step is a fatty acid with a carboxyl group at each end.
|
|
After the three steps, either end of fatty acid can be
attached to coenzyme A. The
molecule can enter the mitochondrion and undergo β oxidation. The final products after successive
oxidation include succinic acid, which
can enter citric acid cycle,
and adipic acid.

Carboxilação do
propionil CoA
Faz-se na presença de ATP pela malonil CoA carboxilase,
enzima necessitando de biotina

Isomerização
A forma D- é isomerizada na forma L pela metilmalonil CoA
recemase

Conversão em sucinil CoA
Faz-se pela acção da metilmalonil- CoA mutase

Carboxilação do
propionil CoA
Faz-se na presença de ATP pela malonil CoA carboxilase,
enzima necessitando de biotina
Isomerização
A forma D- é isomerizada na forma L pela metilmalonil CoA
recemase
Conversão em sucinil CoA
Faz-se pela acção da metilmalonil- CoA mutase
Ácidos não saturados
Os ácidos insaturados têm ligações cis
Os enzimas da beta-oxidação só actuam sobre ligações trans
A transformação cis-trans faz-se através de reacções
sequenciais catalisadas pela dienoilredutase e pela enoil-CoA-isomerase

Capitulo 2
SINTESE DOS ACIDOS GORDOS
O sitio principal para a síntese dos ácidos gordos é o
fígado
Os e.nzimas são diferentes dos do catabolismo
A via sintética
principal implica a polimerização de seis elementos em C2 (acetil-CoA) para dar
um acido em C12(acido palmítico)
A formação do acido palmítico é feita no citoplasma. O seu
alongamento faz-se nas mitocondrias
Sistema citoplasmico
.
Carboxilação do acetil-CoA
A acetil-CoA carboxilase
transfere bicarbonato para o acetil-CoA para formar malonil-CoA


O malonil-CoA é o intermediário
metabólico activo da síntese dos ácidos gordos
A acetil-CoA carboxilase existe
sob duas formas um dimero inactivo e um polímero
activo
O citrato induz a formação do
polímero e o palmitil-CoA provoca a sua despolimerização
Esta reacção requere biotina que está ligada covalentemente
ao grupo amina de um resíduo lisina do enzima
O enzima tem uma forma inactiva fosforilada e uma activa
desfosforilada
Fixação do acetil-CoA
Proteína portadora de acilos(ACP)
Um complexo enzimático catalisa
varias reacções que conduzirão à formação de um acil-CoA com mais dois carbonos
<

http://www.biologie.uni-hamburg.de/b-online/e19/19i.htm<

Este complexo está associado pelo seu fosfato à proteína portadora de acilos (ACP)
A ACP possui fosfopanteina como grupo prostetico, idêntica à que se encontra no CoA
Os acilos ligam-se covalentemente ao grupo sulfidrilo terminal
Reacções envolvidas
· 1ª transacetilação
· Transmalonilação
· Condensação
· 1ª redução
· Desidratação
· 2ª redução
· 2ª transacetilação
1ª transacetilação
A transacilase catalisa a conversão do acetil-CoA em acetil-ACP
Transmalonilação
A transmalonilase transfere o malonil- CoA para o ACP formando acetilmalonilenzima
Condensação
O enzima condensante ou 3-cetoacilsintase catalisa a descarboxilação seguida da transferência do acilo do tiol periférico para o central
Forma-se o beta- cetoacilenzima
1ª redução
A cetoacil-ACP-redutase hidrogena o cetoacilo em hidroxiacilo
Os hidrogénios são cedidos pelo NADPH
Desidratação
A cetoacil-ACP deidrase remove agua entre os carbonos alfa e beta dando um acilenzima insaturado
2ª redução
A enoil-ACP-redutase hidrogena o ACP-acilenzima insaturado num acil-ACP com mais dois carbonos
Os hidrogénios são cedidos pelo NADPH
2ª transacetilação
É transferido um novo radical para para o malonil – coa que se repetirá sucessivamente até se formar palmitil-CoA
Step
|
Enzyme
|
Reaction
|
Description
|
(a)
|
Activates acetyl CoA for reaction with malonyl-ACP
| ||
(b)
|
Activates malonyl CoA for reaction with acetyl-ACP
| ||
(c)
|  |
Reacts priming acetyl-ACP with chain-extending malonyl-ACP.
| |
(d)
|
Reduces the carbon 3 ketone to a hydroxyl group
| ||
(e)
|
Removes water
| ||
(f)
|
Reduces the C3-C4 double bond.
|
Abbreviations: ACP - Acyl carrier protein, CoA - Coenzyme A, NADP - Nicotinamide adenine dinucleotide phosphate.
Regulação
Os mecanismos de regulação impedem que a síntese e o catabolismo se possam fazer simultaneamente
O enzima chave é a acetilCoA carboxilase
Disponibilidade de acetil CoA
O acetil CoA provem do piruvato e do metabolismo de alguns aminoacidos
Disponibilidade de NADPH
Forma-se no ciclo de Dickens-Horecker e na síntese extramitocondrial do isocitrato
Acumulação de acilCoA
de cadeia longa, nomeadamente de palmitil-CoA
Esta situação surge quando por má
metabolização da glicose não se forma glicerofosfato necessário para a síntese
dos trigliceridos
Relações com o
metabolismo da glicose
Todos estes factores estão relacionados com o metabolismo da
glicose
Elongação nas
mitocôndrias
Faz-se pela incorporação de acetil-CoA


Travessia do acetil-CoA
Atravessa as mitocondrias combinado com a carnitina ou
transformando-se em citrato
Origem do NADPH
Descarboxilação
oxidativa do malato
O enzima malico ou malico deidrogenase, enzima citoplasmico,
isoenzima da malato deidrogenase mitocondrial, descarboxila oxidativamente o
malato, \com dformação de NADPH
Descarboxulação
oxidativa do isocitrato
A isocitrato deidrogenase citoplasmica converte o isocitrato
em alfa-cetoglutarato (Fig 129.10)
Ciclo de Dickens-Horecker
É o maior fornecedor
de NADPH
Conclusão
Todas estas reacções estão
associadas ao metabolismo da glicose
Síntese dos ácidos alcool
Formam-se por alfa ou ómega-oxidação
Síntese dos insaturados
Faz-se por dessaturações e elongações sucessivas
Dessaturações
As dessaturases são oxidases utilizando NADP ou NAD,
citocromo P-450 e oxigénio molecular
Os omnivoros e carvivoros só dessaturam entre o C9 e o carbono terminal
Elongação
Faz-se pelo acréscimo de unidades de dois carbonos (acetil
CoA)



Uma outra via é a via dos monogliceridos em que duas moleculasde acil – CoA se combinam com um

http://en.wikipedia.org/wiki/Ketogenesis

Dessaturações
As dessaturases são oxidases utilizando NADP ou NAD,
citocromo P-450 e oxigénio molecular
Os omnivoros e carvivoros só dessaturam entre o C9 e o carbono terminal
Elongação
Faz-se pelo acréscimo de unidades de dois carbonos (acetil
CoA)


Monoinsaturados
Formam-se pela acção de uma delta - 9 -dessaturase sobre o
acilo do acido gordo correspondente( esteárico no caso do acido oleico)
Poliinsaturados
A síntese faz-se por uma série
sucessiva de dessaturações e elongações
Ácidos essenciais
Os ácidos linoleico e linolenico não podem ser sintetizados
O acido linoleico ingerido pode transformar-se em linolenico
pela acção de uma dessaturase
Regulação
A delta 6-dessaturase
é o enzima limitante
É activada por proteínas e pela insulina
É inibida por glucidos,
adrenalina,glucagina e jejum
A sua actividade diminui na
terceira idade, insuficiência hepática e diabetes
Capitulo 3
PATOLOGIA DOS ACIDOS GORDOS
Defeitos da beta-oxidação mitocondrial
Deficiência da acil-CoA deidrogenase de cadeia media(MCAD)
É a mais frequente
Manifesta-se por episodios agudos nos 2-3 primeiros anos de
vida, desencadeados pelo jejum
Os sinais mais importantes são vómitos e letargia evoluindo
rapidamente para coma, convulsões ou colapso cardiorespiratorio
A hipoglicemia está quase sempre presente
A baixa de corpos cetonicos na urina, associada a
hipoglicemia é um sinal característico em todos estes defeitos
A fase aguda trata-se com uma infusão intravenosa de
dextrose para suprimir a lipolise
A alimentação deve
ser ajustada para que durante a noite não haja períodos de jejum superiores a
10 horas
Para mais detalhes consultar
Deficiência em
acil-CoA deidrogenase de cadeia longa(LCAD) ou muito longa (VLCAD)
Não há deficiências isoladas em LCAD
Há alterações graves dos músculos esqueléticos e cardíaco
Nos ataques agudos pode surgir miocardiopatia
Devem-se evitar jejuns de mais de 10 horas
Deficiência em
acil-CoA deidrogenase de cadeia curta(SCAD)
Os sintomas parecem ser devidos à acumulação de metabolitos
dos ácidos gordos de cadeia curta
O sinal mais predominante é a miopatia esquelética
A maioria do doente tem sinais neurologicos e alguns acidose
metabólica severa
O tratamento limita-se a reduzir os periodos de jejum
Deficiência em
hidroxiacildeidrogenase
Ataques de hipoglicemia semelhante aos do MCAD
Os metabolitos tóxicos acumulados podem produzir
retinopatia, insuficiencia hepática progressiva, neuropatia periférica e
rabdomiolise
Os metabolitos podem produzir fígado gordo na mãe
O tratamento implica
a prevenção do jejum
Devem-se restringir os ácidos gordos de cadeia longa e
administrar trigliceridos de cadeia media
O tratamento não
melhora todos os sintomas
Defeitos na alfa-oxidação( Doença de Refsum)
Causas
É um defeito da a-
oxidação do acido fitanico
O ácido fitanico provem do metabolismo do fitol da clorofila
O fitol transforma-se em acido fitanico, que em seguida
seria destruído por alfa-oxidação
Na falta do enzima, o acido fitanico acumula-se
Sintomas
Retinopatia
Polineuropatia
Ataxia cerebelosa
Tratamento
O acido fitanico está presente nos vegetais e na gordura de
ruminantes
O tratamento consiste na supressão de alimentos e outros
produtos contendo clorofila
Defeitos do ciclo da carnitina
Defeito do
transportador
Falta o transportador
de carnitina da membrana que mantem na celula concentrações elevadas de
carnitina
O sintoma mais frequente é a miocardiopatia progressiva
Trata-se com doses farmacológicas de carnitina oral
Deficiência em
carnitina palmitoiltransferase 1
.Hipoglicemia com corpos cetonicos diminuídos em jejum
Provas de função hepatica anormal
Carnitina muito elevada
O tratamento é o mesmo da MCAD
Deficiência em
carnitina-aciltranslocase
Os ácidos gordos não entram nas mitocondrias
Ataques de hipoglicemia e colapso cardiorespiratorio em
jejum
Miocardiopatia e fraqueza muscular
O tratamento é o mesmo da MCAD
Deficiência em
carnitina palmitoiltransferase 2
Na forma neonatal é mortal
Uma forma mais leve manifesta-se no adulto como uma forma de
rabdomiolise episódica
As concentrações sericas de creatinacinase são muito
elevadas
Capitulo 4
METABOLISMO DOS EICOSANOIDES
A síntese dos eicosanoides é feita a partir do acido
araquidonico
Libertação do ácido
araquidonico
O acido araquidonico formado encontra-se nos fosfolipidos
É libertado pela acção das fosfolipase A2
A fosfolipase é
activada por diversos estímulos como a bradicinina, adrenalina e trombina
Via cíclica
Através desta via sintetizam-se as prostaglandinas,
prostaciclinas e tromboxano
PGH2 Sintase(PGS)
Este enzima também é conhecido
como prostaglandina G/H sintase e postaglandina endoperoxido sintase.
Este enzima possui duas
actividades – cicloxigenase (COX) e peroxidase
Há duas formas de actividade COX – COX-1 e COX-2
A COX-1 encontra-se na mucosa gástrica, rim, plaquetas e células endoteliais
A COX-2 é inductivel nos monocitos e macrofagos em resposta
à inflamação
Os factores desencadeantes são o PAF (Platelet Activating
Factor) e a IL-1 (InterLeucina 1)
Tanto a COX-1 como a COX-2 catalisam a conversão do acido
araquidonico em PH2


cortesia de Joyce Diwan
Os anti-inflamatoriios não esteroides inibem este enzima,
acetilado a serina do enzima, o que impede a fixação do acido araquidonico


cortesia de Joyce
Diwan
Formação de
prostaglandinas, prostaciclinas e tromboxano
Formam-se a partir do PH2
Via linear
Formação de HPETE
O acido araquidonico é convertido
em vários ácidoshidroperoxidoeicosapentanoicos (HPETE) por 5-, 12 e 15
lipoxigenases, formando-se os 5,12 ou 15- HPETE

cortesia de Joyce Diwan
Os produtos derivados dos 12 ou
15 HPETE, as hepoxinas e lipoxinas, têm actividades pouco conhecidas
Formação de leucotrienos
O 5-HPETE é convertido em
peptidoleucotrienos
Forma-se primeiro um péptido instável, o leucotrioeno A4
(LTA4)
A glutatião-S-transferase
transfere o glutatião para formar o LTC4
A gama glutamiltransferase remove o acido glutamico
formando-se o LTE4
A glicina é removida por uma dipeptidase para dar o LTB4
Os LTC4, LTD4 e LTDE4
são designados por péptido leucotrienos
Na figura seguinte estão sumarizadas as duas vias

Capitulo 5
METABOLISMO DOS TRIGLICERIDOS
Síntese
Activação do glicerol
A forma activa do glicerol é o glicerofosfato
No fígado, rim e glândula mamaria em lactação a glicerocinase
fosforila o glicerol
No musculo e tecido adiposo, que não têm glicerocinase, a
glicerofosfatodeidrogenase transforma a fosfodihidroxiacetona em glicerofosfato

Activação dos ácidos
gordos

{kind=link}
Uma outra via é a via dos monogliceridos em que duas moleculasde acil – CoA se combinam com um
Capitulo 6
CETOGENESE
No fígado uma porção importante de acetil-CoA pode-se
transformar em corpos cetonicos
Os corpos cetonicos são o acido acético, a acetona e o acido
betra-hidroxibutirico

{kind=link}
Eta.pas
Beta-oxidação
Forma-se acetoacetil-CoA
Inversão da reacção da
tiolase
2
Ac.CoA ------------------ AcAc CoA
Por intermédio do
hidroximetilglutarilCoA
{kind=link}
É uma via comum à síntese do colesterol

Acido
beta-hidroxibutirico e acetona
A b-hidroxibutirico
deidrogenase reduz o acetato em b-hidroxibutirico
A acetona forma-se por descarboxilação espontânea do acido
acetoacetico
Acido
beta-hidroxibutirico e acetona
A b-hidroxibutirico
deidrogenase reduz o acetato em b-hidroxibutirico
A acetona forma-se por descarboxilação espontânea do acido
acetoacetico
Formação a partir de aminoácidos
Há aminoácidos cetogenicos
Destino dos corpos
cetonicos
Destruição nos tecidos extra-hepaticos
O acido acetoacetico é catabolisado pela acetoacetato-suciniltransferas
A acetona é carboxilada em acetoacetato
O acido b-hidroxibutirico é desidrogenado em acetoacetato
Balanço energético
A oxidação dos corpos cetonicos forma 24 ATP
Regulação
O aumento dos corpos cetonicos está sempre relacionado com
um aumento dos ácidos gordos livres circulantes
A formação de corpo gordos ao não serem esterificados
encaminham-se para a cetogenese
Esta situação surge na escassez de glicose(jejum) e na sua
má utilização (diabetes
gordos ao não serem esterificados encaminham-se para a
cetogenese
Esta situação surge na escassez de glicose(jejum) e na sua
má utilização (diabetes)
Sem comentários:
Enviar um comentário